
-
News Message
TCRT 和 CART癌症免疫疗法
- by wittx 2020-06-07
作者:知乎用户
链接:https://www.zhihu.com/question/64720360/answer/223524853
来源:知乎
著作权归作者所有。商业转载请联系作者获得授权,非商业转载请注明出处。白细胞:英文名Leukocyte,white blood cell,简称:Leu或WBC),是无色、球形、有核的血细胞。白细胞一般有活跃的移动能力,它们可以从血管内迁移到血管外,或从血管外组织迁移到血管内。因此,白细胞除存在于血液和淋巴中外,也广泛存在于血管、淋巴管以外的组织中。血液中的白细胞共有五种,按照体积从大到小是:单核细胞、嗜酸性粒细胞、嗜中性粒细胞、嗜碱性粒细胞和淋巴细胞。
单核细胞:是血液中最大的血细胞。是巨噬细胞的前身,具有明显的变形运动,能吞噬、清除受伤、衰老的细胞及其碎片。单核细胞还参与免疫反应,在吞噬抗原后将所携带的抗原决定簇转交给淋巴细胞,诱导淋巴细胞的特异性免疫反应。单核细胞也是对付细胞内致病细菌和寄生虫的主要细胞防卫系统,还具有识别和杀伤肿瘤细胞的能力。巨噬细胞属于单核细胞的一种,具有吞噬作用,所以形体较大。其主要在固有免疫中起作用,主要是吞噬抗原、抗原的处理和递呈。
嗜酸性粒细胞:嗜酸性粒细胞具有粗大的嗜酸性颗粒,颗粒内含有过氧化物酶和酸性磷酸酶。嗜酸性粒细胞具有趋化性,能吞噬抗原抗体复合物,减轻其对机体的损害,并能对抗组织胺等致炎因子的作用。
中性粒细胞:中性粒细胞具有变形运动和吞噬活动的能力,是机体对抗入侵病菌,特别是急性化脓性细菌的最重要的防卫系统。当中性粒细胞数显著减少时,机体发生感染的机会明显增高。
嗜碱性粒细胞:嗜碱性粒细胞中有嗜碱性颗粒,内含组织胺、肝素与5-羟色胺等生物活性物质,在抗原-抗体反应时释放出来。
淋巴细胞:淋巴细胞(lymphocyte)是白细胞的一个类别,是体积最小的白细胞,由淋巴器官产生,是机体免疫应答功能的重要细胞成分。淋巴细胞是一类具有免疫识别功能的细胞系,按其发生迁移、表面分子和功能的不同,可分为T淋巴细胞(简称T细胞)、B淋巴细胞(简称B细胞)和自然杀伤细胞(简称NK细胞)。T细胞和B细胞都是抗原特异性淋巴细胞,它们的最初来源是相同的,都来自造血组织中的干细胞。T淋巴细胞随血循环到胸腺,在胸腺激素等的作用下成熟,而B细胞是在骨髓中分化成熟的。
当受抗原刺激后,T淋巴细胞即转化为淋巴母细胞,再分化为致敏T淋巴细胞,参与细胞免疫,其免疫功能主要是抗胞内感染、瘤细胞与异体细胞等;而B淋巴细胞是先转化为浆母细胞,再分化为浆细胞,产生并分泌免疫球蛋白(抗体),参与体液免疫,其功能是产生抗体,提呈抗原,以及分泌细胞内因子参与免疫调节;NK细胞不依赖抗原刺激而自发地发挥细胞毒效应,具有杀伤靶细胞的作用。
流式细胞分析仪(FCM)根据淋巴细胞表面抗原标志物的不同来检测各淋巴细胞亚群:
主要包括T淋巴细胞(CD3+),B淋巴细胞(CD19+),NK细胞(CD16+CD56+),其中T淋巴细胞可进一步测定辅助/诱导T淋巴细胞(CD3+CD4+)、抑制/细胞毒T淋巴细胞(CD3+CD8+)、CD4+T细胞纯真亚群(CD4+CD45RA+/ CD4+CD45RA+62L+)和记忆亚群(CD4+CD45RA-/ CD4+CD45RO+)、功能亚群(CD28+)、激活亚群(CD38+、HLA-DR+)、凋亡亚群(CD95+)等。
T细胞:T淋巴细胞(T lymphocyte),简称T细胞,属于淋巴细胞的一个分类。T淋巴细胞来源于骨髓的多能干细胞,在胸腺激素的诱导下分化成熟,成为具有免疫活性的T细胞。成熟的T细胞经血流至外周免疫器官的胸腺依赖区定居,并可经淋巴管、外周血等再循环,发挥细胞免疫功能。T细胞的再循环有利于广泛接触进入体内的抗原物质,长期保持免疫记忆。T细胞的细胞膜上有许多不同的标志,主要是表面抗原和表面受体。这些表面标志都是结合在细胞膜上的巨蛋白分子。
T细胞是相当复杂的不均一体、又不断在体内更新、在同一时间可以存在不同发育阶段或功能的亚群,但分类原则和命名比较混乱,尚未统一。按免疫应答中的功能不同,可将T细胞分成若干亚群,一致公认的和常用的有:
细胞毒T细胞(cytotoxic T cell, Tc):消灭受感染的细胞。这些细胞的功能就像一个“杀手”或细胞毒素,因为它们可以对产生特殊抗原反应的目标细胞进行杀灭。细胞毒T细胞的主要表面标志是CD8,也被称为杀手T细胞。
辅助T细胞(helper T cell, Th)在免疫反应中扮演中间过程的角色:它可以增生扩散来激活其它类型的产生直接免疫反应的免疫细胞。辅助T细胞的主要表面标志是CD4,调控或“辅助”其它淋巴细胞发挥功能。
调节/抑制T细胞(regulatory/suppressor T cell, Tr):负责调节机体免疫反应。通常起着维持自身耐受和避免免疫反应过度损伤机体的重要作用。调节/抑制T细胞有很多种,目前研究最活跃的是CD25+CD4+T细胞。
记忆T细胞(memory T cell, Tm):在再次免疫应答中起重要作用。有记忆特异性抗原刺激的作用。T细胞在体内存活的时间可数月至数年。其记忆细胞存活的时间则更长。
TCR:T cell receptor,T细胞(抗原)受体。 TCR为所有T细胞表面的特征性标志,以非共价键与CD3结合,形成TCR—CD3复合物。TCR的作用是识别抗原。由两条不同肽链构成的异二聚体,由α、β两条肽链组成,每条肽链又可分为可变区(V区),恒定区(C区),跨膜区和胞质区等几部分;其特点是胞质区很短。TCR分子属于免疫球蛋白超家族,其抗原特异性存在于V区;V区(Vα、Vβ)又各有三个高变区CDR1、CDR2、CDR3,其中以CDR3变异最大,直接决定了TCR的抗原结合特异性。TCR分为两类:TCR1和TCR2;TCR1由γ和δ两条链组成,TCR2由α和β两条链组成。外周血中,90%-95%的T细胞表达TCR2;而且任一T细胞只表达TCR2和TCR1之一。
CAR-T:嵌合抗原受体T细胞技术,CAR-T(Chimeric Antibody Receptor Engineered T Cell)。
TCR-T:T细胞受体(TCR)嵌合型T细胞技术。
肿瘤免疫治疗,实际上分为两大类。一种把肿瘤的特征“告诉”免疫细胞,让它们去定位,并造成杀伤;另一种是解除肿瘤对免疫的耐受/屏蔽作用,让免疫细胞重新认识肿瘤细胞,对肿瘤产生攻击(一般来说,肿瘤细胞会巧妙伪装,逃脱免疫的监视)。
第一种情况,因为要利用机体自身的免疫细胞,因此,目前多为免疫细胞治疗;后一种主要是阻断被屏蔽的免疫信号,因此,多为小分子药物和抗体药物,如大家熟知的PD-1/PD-L1一类,当然,还有其它类别,甚至联合应用效果更好
而在第一种情况(免疫细胞治疗)中,一直是研究的热点。包括LAK,DC,CIK,DC-CIK,CAR-T,TCR-T,NK,CAR-NK,肿瘤浸润性淋巴细胞(TILs)等等。但是经过十数年的研究,大浪淘沙始到金,眼下有出色作用的是CAR-T,TCR-T这两种细胞治疗方法。而LAK,DC,CIK,DC-CIK基本被证明是无效的,或基本无效,虽然仍然有一些学者在艰苦研究DC相关的治疗策略。而NK,CAR-NK,TIL等技术还有待成熟,有待进一步完善。
CAR-T与TCR-T介绍
作者:知乎用户
链接:https://www.zhihu.com/question/64720360/answer/223524853
来源:知乎
著作权归作者所有。商业转载请联系作者获得授权,非商业转载请注明出处。TCR-T 和 CAR-T这两种技术的一个共同点在于,通过基因改造的手段提高T细胞受体对特异性癌症细胞抗原的识别能力和进攻能力。因此也都统称为“T细胞受体重新定向”技术(T cell receptor redirection)。但两者所使用的方法是不同的。
嵌合抗原受体T细胞技术(CAR-T)以及T细胞受体(TCR)嵌合型T细胞技术(TCR-T)作为当前过继性细胞回输治疗ACT技术的两大最新的免疫细胞技术,因其能够表达特异性受体靶向识别特异性的细胞如肿瘤细胞,受到广泛的关注和研究,从最开始的基础免疫研究转变为临床应用。
基于合成生物学,免疫学,遗传改造技术,使得合成改造的特异性功能加强版的T细胞成为可能。CD19抗原特异性CAR-T细胞用于治疗B细胞白血病白血病和淋巴瘤临床试验中,显示出持续的疾病缓解效果。由于CAR-T/TCR-T技术的优越表现,以及广阔的应用前景,从而使其进入了当前激烈的制药行业竞技舞台中与传统的制药业一较高低。

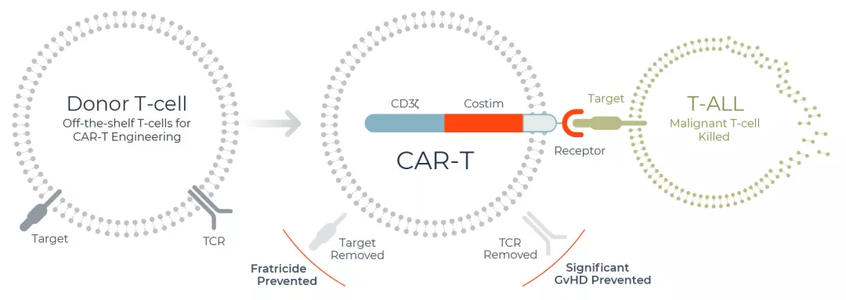
图1:目前细胞治疗的几种途径。通过修饰,使正常供体细胞的同种异体排异反应失活,同时使用抗肿瘤的CAR或TCR武装它;或利用抗肿瘤分子武装患者自体细胞。在实体肿瘤中,活检标本可用来分离TILs并进行扩增。大多数情况下,病人在接受抗肿瘤淋巴细胞输注前需要接受必要的预处理,并须谨慎处置治疗引发的毒性。
T细胞和T细胞受体
T细胞是我们肌体细胞免疫机制中非常重要的一类细胞。这类细胞之所以被称为T细胞是因为它们成熟和分化的过程是在胸腺(Thymus)里完成的。T细胞有一点像我们身体的“纪委巡视组”。它们在肌体内巡逻时,不断地和各种细胞进行“亲密”接触,以检查细胞内部是否正常。而这种接触和检查工作主要是通过一种称之为“T细胞受体”来进行的。
细胞方面和“T细胞受体”交接的是一类被称为“MHC”(或者在人类被称为HLA)的表面蛋白。这类蛋白的功能就是把细胞内部蛋白分解后的片段呈现给T细胞。供T细胞检查。如果T-细胞发现其中有变异的蛋白片段,就可能对细胞发起进攻,摧毁变异的细胞。这也就是我们免疫系统如何能够长期保持我们无癌生活的主要原因。
但癌症还是会经常会发生。这里的原因固然很多,也很复杂。但总体来讲,癌症发生说明我们的T细胞已经失去了识别癌细胞以及摧毁癌细胞的能力。而提高T细胞的识别能力的关键就在于改进“T细胞受体”。从这个角度去思考,就产生了我们刚刚提到的这两种细胞免疫治疗技术:CAR-T和基因修改的T细胞受体。
CAR-T与TCR-T的区别
我们可以打一个简单的比喻。CAR-T是一种“换头”技术。它把TCR的“头”直接换成一个特异性的抗体。这样就可以让T细胞就在抗体的指引下直接进攻癌细胞。和CAR-T这种大手笔的改动相比,基因修改的T细胞受体TCR可以说只是简单地“理了理发”。
用来“理发”的T-细胞并非任何T细胞。这类T细胞通常会“侵入”癌组织。这说明他们对癌细胞有一定的识别能力。事实上,我们可以在这类T细胞中发现对癌细胞相关抗原(Tumo Associated Antigen, TAA)具有特异性识别能力的T细胞。这类抗原包括CEA,Her-2,CD19,gp100, MART-1, MAGA-A3, NY-ESO-1, 等等。这些TAA在不同的癌症中都有相对特殊性的表达。因而也成为免疫系统的攻击对象。
尽管如此,这些天然(或者通过其他手段制造的)的“抗癌”T细胞的识别能力常常较弱,因此不能形成对癌细胞的有利攻击。在这种情况下,可以通过部分基因修改的方法来提高这些TCR对相应TAA的“亲和力”和战斗力。“基因修改的TCR”技术也因此被称为“亲和力增强的TCR”技术(Affinity-EnhanceTCR)。
这次《自然-医学》杂志报道的就是由美国马里兰大学医学院,滨州大学医学院和目前免疫治疗的“当红炸子鸡”之一:Adaptimmune公司联合研发的一款“基因修改的TCR”。在修改了几个关键氨基酸以后,这些基因修改的TCR大大提高了和一种常见的癌症TAA,NY-ESO-1,的亲和力。从而可以用来进攻有NY-ESO-1过量表达的癌症,比如多发性骨髓瘤(MultiplMyeloma)。
在这次临床试验中,80%的多发性骨髓瘤患者有了很好的临床应答。其中70%的患者达到完全或接近完全应答。平均无进展生存期达到了19个月。这样的结果可以说是非常惊人振奋的。
目前这两种方法,发展都比较迅速,但是都不完美。
开发最佳的CAR设计
过去的20年中CAR的设计已经取得了长足的进展(图2)。1991年有3个实验室报道了第一代CAR的设计。Kuwana等人最先构造了嵌合受体,使T细胞不依赖于MHC-I类分子识别靶细胞。Roberts、Finney和Lawson 率先构造了含有CD28或4-1BB共刺激分子的二代CAR。在临床前模型中,使用基于CD28的体外扩增体系,在异种移植瘤模型中4-1BB比CD28对CAR的功效更有益。两种CAR模型都显示出临床前和临床疗效。Long等人的最近研究表明,基于CD28构造的CAR加强并加速T细胞衰竭,而基于4-1BB构造的CAR减缓了T细胞衰竭。我们的研究结果与其结论一致,并且发现CD28内切域能传递使效应T细胞终末分化的信号。从这些研究中得出的周密的结论并不是说某个人源内切域必然比另一个好,而是说CD28信号是生产有效的临床T细胞制品的关键,而4-1BB提升了CAR-T细胞的持久性。

图2:CAR-T细胞的设计。第一代CAR引入了CD3ζ链或类似的信号域。基于抗体再定向的T细胞首先由Kuwana设计并由Eshhar完善。Roberts和Finney首先设计了整合CD28或CD137信号域的第二代CARs。
为了进一步改良CAR的设计,许多研究组开始着眼于发展第三代CAR,不仅包括“信号1”、“信号2”,还包括了额外的共刺激信号。不同研究者们用不同材料开展的研究所得到的第二代CAR和第三代CAR的比较结果是矛盾的。不过,新一代CAR-T能有效控制毒性。
透视ACT毒性
十多年来,许多ACT临床试验表明各类工程化的细胞产品相当安全,但有效性相对缺乏。过去的4年里,ACT的疗效发生了一场革命性变化,体现在工程化的T细胞可以在体内大量扩增,甚至在某些情况下可以持续性存活。这彻底的改变了ACT的疗效,尤其体现在CAR-T细胞应用于癌症治疗,但是在显示出疗效的同时,伴随有毒性。
细胞因子释放综合征(CRS)
CAR-T治疗骨髓源肿瘤最显著的毒性是CRS。高度增殖的T细胞能引起CRS,表现为高热和肌痛,不稳定的低血压和呼吸衰竭。这是一个意想不到的结果,因为在临床前动物模型中没有出现类似症状。从CRS观察中发现一个关键的点,除了预期的效应细胞因子INF-γ外,IL-6在CART治疗的细胞指数级增殖期间也会迅速提升。CRS可能直接与另一个毒性相关联,即巨噬细胞活化综合征。幸运的是,针对这些严重的CRS,寻找到了一种治疗方法,应用IL-6受体拮抗剂tocilizumab来阻断IL-6的作用。
采用高度增殖的CART治疗ALL患者,确认了IL-6的出现和tocilizumab对严重CRS的疗效,并且我们观察到了双特异性T细胞衔接抗体blinatumomab引起的严重CRS具有类似机制。
另一个重要发现是,严重的CRS几乎发生在高疾病负荷病人身上。这意味着将CAR应用于疾病早期患者身上,在疾病恶化之前使用CART来杀灭肿瘤,发生严重CRS风险将大大降低。
关于靶向毒性
T细胞增殖可直接导致CRS。它不依赖于改造的T细胞的抗原特异性,即使CAR是相同的,治疗不同的疾病有可能CRS风险和症状也会有所不同。例如在治疗CLL和弥漫大B时CRS较轻而治疗ALL时CRS相对较重。
除了CRS,还存在由工程改造的T细胞的抗原特异性导致的“靶向”毒性。例如溶瘤综合征,它直接是由肿瘤细胞的裂解而导致的。当CARs靶向于B细胞表面表达的靶点如CD19时,会引起B细胞发育不良,这就是一个“靶向”毒性,但却错误的攻击了正常组织细胞的结果。只要CD19 CAR-T细胞长时间存在,B细胞发育不良的情况就不会改善,因此,B细胞缺乏可作为CAR-T疗效持续的标志。B细胞发育不良与CD20特异性单抗治疗一样会造成严重的低丙种球蛋白血症,需要静脉注射免疫球蛋白。不像CD20单抗rituximab引起的相对较轻的低丙种球蛋白血症,CAR引起的长期B细胞发育不良,静脉注射免疫球蛋白非常必要。
最近报告了输注改造的T细胞引发致命毒性的2个案例,有一例患者接受了HER2-CAR治疗,两例患者接受了靶向MAGE-A3的TCR-T细胞治疗。在这2个案例中,均是因为正常组织表达这些靶点,导致急性不可逆的心肺毒性。所有的靶向毒性均是由于改造的T细胞无法区别表达靶向抗原的正常细胞和肿瘤细胞所致。
高亲和力TCR改造的T细胞引起的毒性可能与亲和力成熟过程有关,因为“非肿瘤靶向”的高亲和力TCR的产生在生理条件下将受到限制。在设计TCR的过程中,小心筛选高亲和力的TCRs,并研究其对靶蛋白之外的其他抗原表位的识别显得十分必要。根据已有报道,输注HLA-A2/MAGE-A3特异性的TCR-T细胞能够产生神经毒性。
神经毒性
CAR-T治疗白血病会引起神经系统症状,这是一个预期之外且目前尚不清楚的现象。几个研究小组报道,这些症状具有多样性但可自行消退,如谵妄、语言障碍、运动障碍、缄默症和癫痫发作。虽然与全身CRS的发生有些时间上的关联,当然也与CAR-T存在于脑脊液中相关,但是使用tocilizumab这些症状不会有改变。这些症状的机制与靶组织仍有待确认
其他毒性
输注活化的T细胞存在引起自身免疫性疾病的风险。靶向黑色素瘤的ACT治疗可诱发白癜风。输注活化的T细胞产生如皮疹、结肠炎、垂体炎等毒性的报道较少。CAR修饰的T细胞尚未出现这些问题。输注同种异体T细胞存在抗宿主移植病的潜在风险。这可能会引起曾接受同种异体造血干细胞移植的患者的担心。幸运的是,从受体采集已耐受供体来源的T细胞(而不是捐献者),抗移植物宿主病的风险会非常低。当然有一种方法是利用自杀系统来限制毒性,如经典的caspase-9
诱导表达系统,或包含特定表面靶点如CD20。在含有caspase-9诱导系统下,一种水溶性活化剂可激活Caspase系统导致T细胞凋亡。包含CD20基因修饰的CAR-T细胞,可用单抗如rituximab来清除“违规”的细胞。该领域面临的问题
近期的各种实验室基础研究,正在解决科学与工程方面遇到许多挑战。下面我们谈一谈当前面临的挑战。
细胞制品的构成
早期的细胞治疗主要是输注高分化的CD8+T细胞。这些CTL细胞具有很强的细胞毒性,但是输注后没有足够的复制能力,除了个别情况外,注入患者体内的T细胞持久性较差。目前普遍同时输注CD4+与CD8+T细胞,很可能是因为CD4+T细胞提供生长因子和其他信号以维持输注的CTL的功能和活性。
另外,在小鼠试验中的研究有时候会误导处理人源T细胞的方法。例如,由于受到端粒退化的影响,人源T细胞的复制能力有限,在小鼠模型中却没有这个特点,而人CD4+细胞比小鼠CD4+细胞具有更强的细胞毒性。
现在的问题是是否需要利用流式细胞技术或其他的方法分离细胞亚群,并进行独立培养。CD4和CD8+T细胞最佳细胞培养条件不同,这因为CD +和CD8 +T细胞信号转导途径不一样。此外,细胞分离法可去除对效应T细胞疗法存在潜在危害的Treg细胞。况且,需要从输注的T细胞中去除肿瘤细胞,对白血病患者而言,这尤其具有挑战性。然而,在GMP生产中有些技术使细胞制备成本大大增加,如通过流式细胞术或磁珠分选细胞可使成本迅速增加10000美元甚至更多。
年龄问题
与上述问题相关,输注处于什么分化阶段的T细胞最佳?对小鼠和人类的研究结果表明,naive或中心记忆细胞是最好的。对只有少量naive T细胞的老年人而言,如何获取这类细胞是个挑战,且化疗或其他疾病通常会进一步放大难度。从我们采用CD19特异性CAR-T治疗白血病患者的数据中表明,扩增能力是预测成功的最重要的生物学标志。
一种方法是在全血淋巴细胞中分离中心记忆或naive T细胞。与此相反,一个更简便粗放的方法是使用大量T细胞在特定培养条件下维持naive或中心型记忆细胞的扩增。后一种方法基于特异性共刺激信号可以促使细胞定向分化的原理。如CD28的刺激可使CD4细胞保持中心记忆细胞状态,4-1BB可以促使CD8中心记忆细胞的增殖。相反ICOS共刺激分子可以促使和稳定Th17细胞的生长。利用T细胞的干细胞样特性是非常有希望的,Wnt信号通路的激活可以促进记忆干细胞的增殖。
是否存在统一的最佳细胞制品配方?抑或针对不同的肿瘤需要不同的配方?
CD19 CAR-T治疗B细胞恶性肿瘤显示出的惊人且意想不到的疗效能否出现在治疗实体瘤患者身上还未可知。我们发现CAR-T在体外杀灭低分化腺癌细胞的效率类似于白血病。然而,仍然有待进一步确定是否需要修改细胞组成类型或细胞培养条件,用来优化实体瘤患者体内改造的T细胞的转运和持久性。与恶性血液疾病的散在分布不同,采用细胞工程的方法优化T细胞向实体瘤块转运,如成胶质细胞瘤和胰腺癌,可能会有所不同。已提出一种方法是利用靶向策略,如采用基因工程的手段,使用各种趋化因子或趋化因子受体促使T细胞向肿瘤部位富集。其他方法包括对肿瘤或患者进行预处理,如进行放疗,注射溶瘤载体以及在瘤内直接注射T细胞。
最优的共激活结构域在CD4+和CD8+ T细胞内是一样的吗?
以往研究表明不同亚群的T细胞培养条件是不同的。这就提出一个问题,不同淋巴细胞亚群的T细胞是否需要不同的信号分子进行武装呢?为着手解决这个问题我们对CD4+和CD8+T细胞胞内区CD28, 4-1BB和ICOS进行了评估。
在异种移植腺癌的人源化小鼠中,我们发现采用ICOS信号域的CD4+ CAR-T细胞更为优越,而CD8+T细胞常优选4-1BB。如此增加细胞制备的复杂度是否能使临床实验受益尚有待进一步确定。此外,最佳的T细胞亚群比例和信号分子的构造在不同的肿瘤微环境中很可能会有差异。比如在一些临床前肿瘤模型中改造的Th17细胞比Th1细胞更优越。
生存与毁灭:自杀结构能否减弱非肿瘤靶向引起的毒性?
使用改造的T细胞最大的不确定性是利用生物合成技术诱导的新的、增强的细胞效应是否会引发意外的脱靶毒性。CAR-T细胞的非肿瘤靶向毒性和TCR细胞的脱靶毒性被格外关注。有多种方法可以缓解这些问题。我们发现利用编码CAR的mRNA转染T细胞,限制CAR仅在转染T细胞中表达,对评估出现的直接毒性有帮助。当出现毒性反应,停止细胞输注,毒性会迅速减弱。研究人员也提出了许多诱导基因修饰T细胞凋亡的方法。这些方法很可能都会被整合,以便按需清除T细胞,满足严格的安全需求
结论
基于正在进行的试验,表达CARS和TCRS的T细胞已经为广泛的商业应用准备就绪。华尔街的资源正推动这个曾因投资不足而被限制的领域的进步。在输注的T细胞中引入或删除基因,提供了能克服肿瘤免疫抑制微环境的新细胞制品,并且最后可能不需要使用免疫检查点封闭抗体。ACT先进的基因工程技术的到来,将会使利用合成生物学原理进行癌症、慢性感染和自身免疫性疾病的治疗取得显著进展。
Share Http URL: http://www.wittx.cn/get_news_message.do?new_id=285

Best Last Month
Information industry by wittx
Information industry by wittx
Information industry by wittxUltrauniform, strong, and ductile 3D-printed titanium alloy through bifunctional alloy design
.jpg)
Information industry by wittx
Information industry by wittx

Medical science by wittx
Information industry by wittx
Information industry by wittx

Information industry by wittx
Information industry by wittx
